ORIGEN y CARACTERÍSTICAS GENERALES DE LA SEMILLA
El desarrollo exitoso de la semilla depende de múltiples influencias en todos
y cada uno de los estados de su formación. Además, su estructura está
estrictamente unida a la función de la misma, por lo tanto, el estudio de sus
características nos permite comprender sus posibilidades futuras de éxito.
En las angiospermas las semillas se desarrollan a partir de los óvulos como
consecuencia de la doble fecundación (uno de los gametos masculinos se une con
la oósfera; el segundo, a los núcleos polares). En dicho momento, el óvulo
consiste de una nucela central, que contiene al saco embrionario y uno o
dos tegumentos. En un óvulo bitégmico la nucela esta rodeada por un
tegumento interno y otro externo generalmente de menor desarrollo. La apertura
delimitada por los extremos de ambos tegumentos forma el micrópilo,
figura 2. Sin embargo, los tegumentos pueden tener distinto desarrollo y
sus extremos presentarse excéntricos, en zig zag, como en los óvulos jóvenes de
la soja (Glycine max) o en el palo borracho (Ceiba insignis). De
esta forma las aperturas se denominan endostoma y exostoma respectivamente. Los
óvulos unitégmicos son los que presentan un solo tegumento, como en las
gimnospermas; en este caso, contienen a la nucela y al prótalo. Excepcionalmente
se presentan óvulos atégmicos (sin tegumentos).
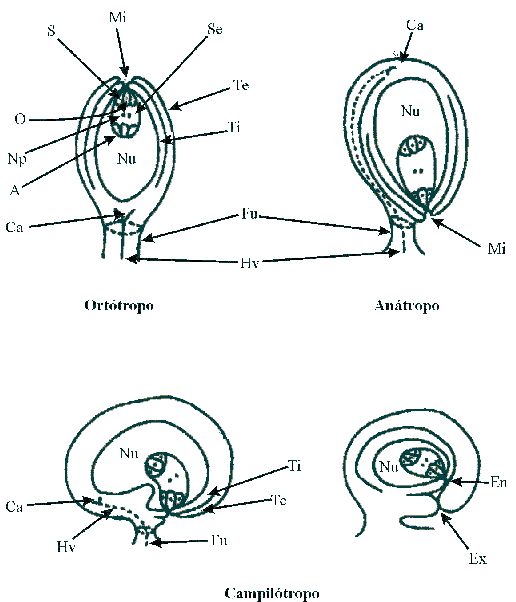 |
| FIGURA 2: Partes del
óvulo y tipos de óvulos. |
Te,
tegumento externo; Ti, tegumento interno; Fu, funículo; Nu, nucela;
Ca, cálaza; Hv, haz vascular; S, sinérgidas; Se, saco embrionario; O,
oósfera;
A, antípodas; Np, núcleos polares; En, endostoma; Ex, Exostoma. |
Los óvulos se unen a la placenta por el funículo, por donde circula el haz
vascular que nutre al óvulo y que se ramifica en la cálaza, ubicada en la base
de la nucela. Generalmente se reconocen tres tipos de óvulos: el ortótropo u
átropo, donde funículo, cálaza, nucela y micrópilo se encuentran en línea recta;
el anátropo, en el cual el óvulo sufre una curvatura o giro de 180° y el
funículo se suelda lateralmente al tegumento externo del óvulo, manteniendo la
nucela recta; y el óvulo campilótropo que presenta la nucela curva, figura 2. Mientras los óvulos anátropos son los
más frecuentes entre la mayoría de las familias de las angiospermas y los
campilótropos se encuentran en importantes familias como las leguminosas
papilionoideas y crucíferas o brasicáceas; los óvulos ortótropos sólo ocurren en
unas pocas familias de angiospermas como las juglandáceas y poligonáceas y son
comunes en las gimnospermas.
que presenta la nucela curva, figura 2. Mientras los óvulos anátropos son los
más frecuentes entre la mayoría de las familias de las angiospermas y los
campilótropos se encuentran en importantes familias como las leguminosas
papilionoideas y crucíferas o brasicáceas; los óvulos ortótropos sólo ocurren en
unas pocas familias de angiospermas como las juglandáceas y poligonáceas y son
comunes en las gimnospermas.
Luego de la fecundación y una vez completado el proceso de embriogénesis, los
tegumentos del óvulo se transforman en episperma o cubierta seminal.
El cuerpo del óvulo, formado por la nucela y el saco embrionario, es ocupado por
el embrión y el tejido que almacena las sustancias de reserva (endosperma,
perisperma o prótalo, según sea el origen de las mismas).
La semilla mantiene algunas características del óvulo del cual deriva.
En los óvulos bitégmicos, generalmente la primina (tegumento externo) se
transforma en la testa; la secundina (tegumento interno), en el tegmen.
Sin embargo, las transformaciones pueden ser variadas y no siempre intervienen
ambos tegumentos en la formación la cubierta seminal. En la
mayoría de las leguminosas como la soja (Glycine max), sólo el tegumento
externo se diferencia en las distintas capas celulares que forman la cubierta
seminal, figura 3.
 |
Figura 3:
Capas celulares de la cubierta seminal de Glyclne max (L.)
Merrill.
A, corte transversal del hilo; B, detalle del recuadro;
Cu, cutícula; Co, cotidelones; Cs, cubierta seminal; Hi,
hilo;
Ma, macrosclereidas; Os, osteosclereidas; Re, restos
de endosperma. |
| Fotografías gentileza
del Ing. Agrón. (Ph.D.) Cristiano Casini |
En otros casos, los tegumentos se reducen y forman una cubierta seminal
delgada como en el maní (Arachis hypogaea) y nogal (Juglans regia),
o adquieren características especiales para favorecer la diseminación; por
ejemplo en las semillas aladas de los pinos (Pinus spp.) o en las del
quebracho blanco (Aspidosperma quebracho-blanco), figura 4.
 |
Figura 4: Semillas
aladas
A, pino (Pinus
sp.); B, quebracho blanco
(Aspidosperma quebracho-blanco Schltdl. ) |
La semilla se separa del funículo por abscisión, dejando en la cubierta
seminal una cicatriz denominada hilo, cuya forma puede variar desde
circular a elíptica. Al mismo tiempo, pueden persistir restos de tejido
funicular y formar un arilo seco o carnoso. El micrópilo, frecuentemente
permanece visible como un poro circular, deltoide (D),
epsiloide (Y), o como una simple hendidura lineal. Sin embargo, a veces no se
advierte por estar obliterado e incluído en el hilo.
En las semillas que derivan de óvulos ortótropos el micrópilo y el hilo se
observan en extremos opuestos, como por ejemplo en la semilla cerebroide del
nogal (Juglans regia) o del pecan (Carya pecan); mientras que en
las derivadas de óvulos anátropos o campilótropos, el hilo y el micrópilo se
encuentran muy próximos. En las semillas provenientes de estos dos últimos tipos
de óvulos, la parte del funículo que gira y se suelda al tegumento externo forma
un reborde longitudinal que se denomina rafe, delimitado entre el hilo y
la base del óvulo (cálaza) y entre ésta y el micrópilo se define el antirafe,
figura 5.
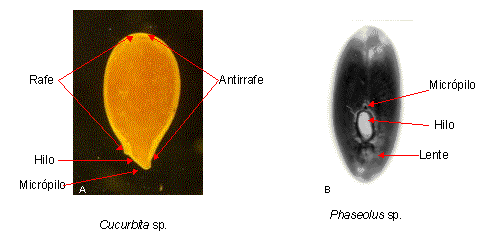 |
Figura 5: Semillas derivadas de
distintos tipos de óvulos
A, óvulo anátropo (Cucurbita sp.); B, óvulo campilótropo (Phaseolus
sp.) |
Ambas zonas son aproximadamente simétricas si derivan de un óvulo anátropo,
como por ejemplo en los algarrobos (Prosopis spp.) y el timbó (Enterolobium
contortisiliquum); mientras que si la semilla deriva de un óvulo
campilótropo, la rafe es notablemente más corta como en los porotos (Phaseolus
spp.), tréboles (Trifolium spp.), la soja (Glycine max) y arveja (Pisum
sativum).
En síntesis, la semilla botánica se caracteriza por presentar una cubierta de
protección, el episperma o cubierta seminal, derivada de los
cambios experimentados por los tegumentos del óvulo durante la formación
de la semilla; el embrión, resultado del proceso de embriogénesis y el
endosperma, presente en mayor o menor proporción, que es el tejido nutricio
originado por endospermogénesis a partir del núcleo endospérmico.
Por otro lado, cuando las sustancias de reserva provienen de la nucela, el
tejido nutricio es el perisperma. En las gimnospermas el tejido
reservante es el prótalo, formado por sucesivas divisiones mitóticas de
la megáspora antes de la fecundación. Consecuentemente, en las semillas de las
angiospermas, la cubierta seminal representa la generación de la planta madre,
mientras que el embrión y el endosperma constituyen una nueva generación. Cuando
las sustancias de reserva derivan de la nucela, el perisperma también forma
parte de la generación de la planta madre; al igual que la cubierta seminal y el
prótalo en las gimnospermas.
En el fruto-semilla de las gramíneas (poáceas), por fuera del episperma y
soldado a éste, se encuentra el pericarpo, como en los trigos (Triticum
spp.) y el maíz (Zea mays). En las avenas (Avena spp.), el
cariópsis va acompañado de las brácteas (lema y pálea), que son piezas extra
ováricas. En los aquenios de las asteráceas (compuestas) como en la lechuga (Lactuca
sativa) y el girasol (Helianthus annuus), por fuera de la semilla
botánica se encuentra, además del pericarpo, el clamidocarpo (hipanto que rodea
al ovario cuando éste es ínfero). Las características morfológicas de todas las
semillas en sentido amplio, permiten identificar las semillas de distintas
especies.
Desde el punto de vista agronómico, resulta fundamental el conocimiento de
las semillas de dos grandes familias o grupos taxonómicos: las leguminosas o
fabáceas y las gramíneas o poáceas; importantes por en el número de especies
involucradas en los distintos sistemas de producción agropecuaria. Las semillas
de otros grupos, de valor agroalimenticio, como solanáceas (tomate,
Lycopersicon esculentum; pimiento, Capsicum annuum), cucurbitáceas
(calabazas, Cucurbita spp. o melón, Cucumis melo) entre otras,
presentan en términos generales un patrón común a las anteriores con ligeras
variantes.
CUBIERTA SEMINAL
La estructura de la cubierta seminal o episperma guarda una estrecha relación
con las funciones que desempeña: protección, dispersión y absorción de agua. Es
la primera defensa de la semilla contra las condiciones adversas del medio que
la rodea, no sólo la proteje de las tensiones mecánicas y la invasión de
organismos patógenos, sino también de las fluctuaciones de humedad y
temperatura. Por esta razón, la integridad de la cubierta seminal juega un rol
importante en la conservación de la calidad de la semilla.
Poco antes de alcanzar la madurez fisiológica,
la semilla comienza a deshidratarse en la planta madre, primero a través de la
cubierta seminal y en la etapa final continúa la deshidratación por el hilo. En
general, las cubiertas seminales son secas y permeables al agua. A medida que la
semilla madura en la planta madre es común que las capas celulares externas se
transformen en esclereidas como en las leguminosas como por ejemplo en los
tréboles (Trifolium spp., Melilotus spp.) y las vicias (Vicia
spp.). En éstas especies, algunas semillas se vuelven impermeables, y se las
conoce como semillas duras ya que para poder absorber agua durante la
imbibición, necesitan de un tratamiento denominado escarificación, que
remueve un sector de la cubierta seminal.
En pasturas naturales, la presencia de semillas duras constituye una "ventaja
estratégica", ya que de esta forma, el suelo mantiene un banco natural de
germoplasma. Bajo estas condiciones, las semillas se escarifican en forma
natural, por acción microbiana o al ser arrastradas por el viento, agua o por el
pasaje a través del tracto intestinal de los animales que las ingieren. Las
semillas germinan en forma progresiva permitiendo una buena distribución y
cobertura vegetal, determinando una mayor vida útil de la pradera. Sin embargo,
un alto porcentaje de semillas duras se traduce en una desventaja cuando el
productor tiene como objetivo la producción de forraje o semillas, que requieren
de una germinación e implantación pareja y una maduración sincronizada.
La práctica más común cuando se siembran forrajeras tales como la alfalfa (Medicago
sativa), vicias (Vicia spp.) o algunas especies de tréboles del
género Trifolium es la escarificación física, que consiste en hacer girar
las semillas en tambores con papel de lija. Otra alternativa es la
escarificación química, tratando a las semillas con ácido sulfúrico. Una
desventaja de estas prácticas, es que en el escarificado se puede dañar al
embrión de modo tal que determine la pérdida de viabilidad
de las semillas, con todas las implicancias económicas que derivan de esta
situación.
En la actualidad, mediante selección, se ha logrado que muchas de estas
especies con semillas duras hayan perdido esta cualidad y aseguren una
germinación uniforme y rápida de los cultivos, resultando en una madurez pareja
y sin problemas para su cosecha.
La cubierta seminal puede ser lisa y brillante como en el yuyo colorado (Amaranthus
quitensis), rugosa como en el chamico (Datura ferox), la calabacilla
(Silene gallica) y la verdolaga (Portulaca oleracea) o presentar
pelos epidérmicos como los del algodón (Gossypium hirsutum), que son la
fuente del algodón comercial, figura 6.
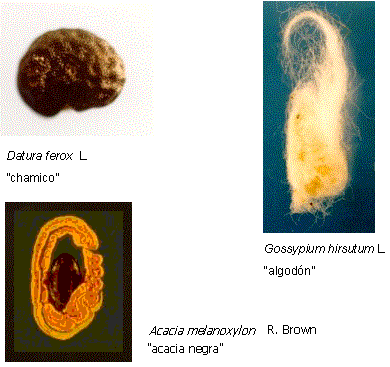 |
| Figura 6: Diferentes
caracterísiticas de la cubierta seminal |
Algunas semillas como las de lino (Linum usitatissimmum) contienen
sustancias muscilaginosas (higroscópicas) en la cubierta que favorecen la
retención de agua durante el proceso de imbibición. Otras semillas presentan
capas carnosas como en la granada (Punica granatum). Al mismo tiempo,
pueden presentar pigmentos y además exhibir excrecencias secas o carnosas más o
menos desarrolladas y comestibles, denominados arilos en sentido amplio y
que resultan atractivos para los animales. Dichos arilos son restos del funículo
que persisten en la superficie de la cubierta y cubren gran parte de la semilla
como en la nuez moscada (Myristica fragans), la acacia negra (Acacia
melanoxylon),o como en la semilla de ricino (Ricinus communis) que
presenta un arilo micropilar denominado carúncula.
Es muy importante tener en cuenta que todas las características que se
encuentran en las semillas y que favorecen su diseminación, lo son tanto para
aquellas especies que nos interesa que se preserven, como para las que son
consideradas malezas en los cultivos y que además pueden dispersar plagas y
enfermedades. Por lo tanto se deben tener muy presentes para el control de las
mismas. Además, su conocimiento permite al productor, la toma de decisiones
respecto a la compra de una determinada calidad de semilla, como así también,
una vez cosechada su propia semilla, evaluar el nivel de contaminación con otras
de malezas, y al reconocerlas, planificar el sistema de limpieza más eficiente
(elección del tipo de maquinaria que le permitirá eliminar aquellas semillas que
contaminan a las de su cosecha). Inclusive puede evaluar y replantear el sistema
de control de las malezas durante las distintas etapas de la producción para la
campaña siguiente. Por ejemplo, una práctica común entre los productores de
semillas de alfalfa (Medicago sativa), cuando detectan algún lote
contaminado con semillas de cuscuta (Cuscuta spp.), es la limpieza del
mismo. Esta práctica postcosecha, consiste en hacer pasar el lote contaminado
por polvo de hierro, que se adhiere a la cubierta seminal rugosa de la cuscuta,
y que posteriormente al pasar por un electroimán queda retenida por el mismo,
dejando limpio el lote.
Si analizamos cualquier semilla del grupo de las leguminosas (alfalfa,
Medicago sativa; soja, Glycine max) veremos que responden a un patrón
común, figura 7.
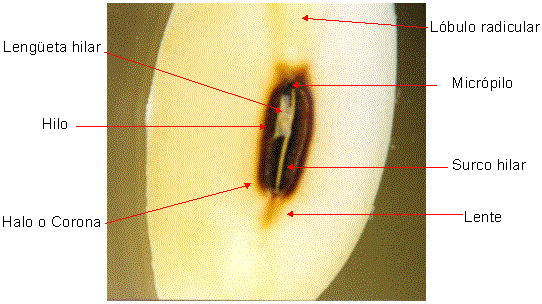 |
| Figura 7: Características de la
región hilar en soja (Glycine max (L.) Merrill) |
En vista externa podremos distinguir, enmarcado por un halo o corona, una
cicatriz más o menos elíptica, el hilo y la lengüeta hilar (arilo
seco), que corresponden al funículo del óvulo que durante el desarrollo de la
semilla lo mantenía unido a la placenta del ovario. El surco hilar divide
al hilo en toda su longitud. Como las especies de este grupo generamentel
derivan de óvulos campilótropos o anátropos, muy próximo al hilo o incluído en
él se encuentra el micrópilo y comúnmente por sobre éste se distingue el
lóbulo radicular. Opuesto a éste se encuentra la lente, que se distingue
como una zona sobre elevada, a veces discolor. En recientes trabajos se ha
establecido la importancia de la lente durante la imbibición de las semillas
denominadas "duras", siendo éste el lugar por donde se produciría la entrada del
agua a una velocidad "controlada" evitando daños en el embrión, que resultarían
en un fracaso del establecimiento de las futuras plántulas. La función del hilo
es la de mantener la humedad de la semilla más o menos constante una vez que
ésta ha alcanzado su madurez. La apertura del surco hilar se produce cuando la
humedad de la semilla es superior a la del aire que la rodea, permitiendo que se
libere vapor de agua. Cuando la humedad relativa externa es superior a la de la
semilla el surco se cierra.
SUSTANCIAS DE RESERVA
En las angiospermas, poco después de la fecundación, comienzan las divisiones
mitóticas tanto del núcleo endospérmico (proceso de endospermogenésis), como del
cigoto (embriogénesis). El crecimiento del embrión, en un principio, es más
lento que el del endosperma, y ambos se desarrollan inicialmente a expensas del
tejido nucelar, que cede sus reservas y cuyas células colapsadas, una vez madura
la semilla, tapizan internamente la cubierta seminal.
El endosperma desempeña una función importante como intermediario, tanto en
la nutrición del embrión durante su desarrollo y maduración, como también en el
crecimiento de la plántula, en la germinación de aquellas semillas
endospermadas. El endosperma ejerce un control hormonal en el crecimiento y
diferenciación del embrión y en su ausencia el embrión generalmente aborta.
Además, Durante mucho tiempo se creyó que el endosperma era un tejido muerto,
sin embargo en la actualidad se sabe que en la mayoría de las familias está
compuesto por células vivas, aunque en algunos casos se combinan células vivas y
muertas como sería en el caso de las gramíneas y leguminosas. El endosperma no
solo nutre al embrión durante los estados tempranos de su crecimiento y
diferenciación (embriogénesis), sino que además puede restringir físicamente la
emergencia de la radícula (germinación) como ocurre en caso del tomate (Lycopersicon
esculentum) y la lechuga (Lactuca sativa).
Según su desarrollo se reconocen tres tipos de endosperma. El más primitivo y
frecuente es el nuclear, en el que primero se suceden las divisiones del
núcleo endospémico, y en una etapa final se produce la formación de las paredes
celulares (citocinesis). El tipo celular deriva del anterior y a cada
división le sigue la formación de la pared, presenta haustorios por
diferenciación celular y es característico de familias más evolucionadas
(gamopétalas) y con semillas pequeñas. El tipo helobial se da en
algunas monocotiledóneas, se caracteriza porque en la primera división se forman
dos células de distinto tamaño, una pequeña denominada basal, que se ubica hacia
la cálaza y una célula micropilar, de mayor tamaño, que origina el endosperma
central.
En la soja (Glycine max) el endosperma, que es de tipo nuclear,
presenta 8 núcleos cuando el proembrión es bicelular. Posteriormente, el
endosperma es consumido por el embrión en desarrollo y sólo permanecen una o dos
capas de células en la semilla madura.
Según la localización y el origen de las sustancias de reservas, las semillas
clasifican en:
- ENDOSPERMADAS: El endosperma, como tejido reservante, ocupa un
volumen importante en la semilla. Su dotación cromosómica (resultado de la
fusión de los nucleos polares de la célula media y una gameta masculina) es
triploide (3n). A su madurez, tiene una consistencia esponjosa y rodea al
embrión. Son ejemplo entre las dicotiledóneas, el pimiento (Capsicum annuum),
tomate (Lycopersicon esculentum), chamico (Datura ferox) y la
mayoría de las monocotiledóneas como el trigo pan (Triticum aestivum),
las avenas (Avena spp.) y la cebolla (Allium cepa).
- EXENDOSPERMADAS*: Cuando las sustancias de reserva se acumulan en
los cotiledones, el máximo desarrollo del embrión se produce a expensas del
endosperma que es digerido y queda reducido a una fina capa de células
parenquimáticas que rodean al embrión. Los cotiledones son muy voluminosos, y
en ellos se encuentran acumuladas las reservas. Son ejemplo la mayoría de las
leguminosas como el poroto (Phaseolus vulgaris), la arveja (Pisum
sativum), incluso las asteráceas como el girasol (Helianthus annuus)
o cucurbitáceas como el zapallo criollo (Cucurbita maxima).
- PERISPERMADAS: Las resevas se acumulan en el perisperma, tejido que
resulta de las numerosas mitosis que se producen en la nucela del óvulo, y por
lo tanto su dotación cromosómica es diploide (2n). Son ejemplo de semillas
perispermadas la acelga (Beta vulgaris var. cicla), pimienta (Piper
nigrum), verdolaga (Portulaca oleracea) y morenita (Kochia
scoparia).
- PROTALADAS: Las sustancias de reserva se acumulan en el prótalo,
que deriva de las numerosas divisiones mitóticas de la megáspora, por
consiguiente es haploide (n) y que resulta en una estructura homóloga al saco
embrionario de las angiospermas, siendo el caso típico de las gimnospermas (Pinus
spp, Cedrus spp. etc.).
* En las semillas exendospermadas generalmente persisten restos de endosperma
formado por un par de capas de células que contienen hemicelulosa como reserva
de hidratos de carbono. La diferenciación entre los tipos de semilla
endospermada y exendospermada no es absoluta. Debido a que hay una tendencia
evolutiva de las espermatófitas hacia las semillas exendospermadas, la presencia
de abundante endosperma que rodea a un pequeño embrión es considerado un
carácter primitivo.
Como se mencionó anteriormente, las semillas de las leguminosas y los
cariópsides de los cereales, constituyen una importante fuente de alimento cuyo
valor radica en el almacenaje de hidratos de carbono, proteínas y lípidos que se
depositan durante el desarrollo y maduración de la semilla. Las cantidades
relativas en porcentaje promedio de diferentes especies se presentan en la
siguiente tabla:
| TABLA 1: |
PORCENTAJE DE HIDRATOS DE CARBONO,
PROTEÍNAS Y LÍPIDOS ALMACENADOS EN SEMILLAS DE DISTINTAS ESPECIES |
| Especie |
HCa |
Proteínas |
Lípidosb |
Almacenamiento |
| trigo |
75 |
12 |
2 |
endosperma |
| maíz |
80 |
10 |
5 |
endosperma |
| arroz |
86 |
10 |
2 |
endosperma |
| girasol |
27 |
13 |
24 |
embrión |
| soja |
26 |
37 |
17 |
cotiledones |
| maní |
12 |
31 |
48 |
cotiledones |
| colza |
6 |
20 |
42 |
embrión |
| a:
principalmente almidón |
| b: en cereales los lípidos se almacenan en el
escutelo |
| modificado de Bewley & Black. 1994.
|
CARBOHIDRATOS
El almidón es el principal representante de los carbohidratos como sustancia
de reserva de la mayoría de las semillas; tanto en las endospermadas como en los
cotiledones de las exendospermadas. Se deposita en foma de gránulos que varían
de 2 a 100 micrómetros de diámetro en los amiloplastos de las células de
reserva. Los granos de almidón están constituídos por capas concéntricas
alternativamente brillantes y mate que se depositan alrededor de un punto
denominado hilo. Cada capa representa la cantidad de almidón formada en 24 horas
y en ellas se distinguen claramente dos zonas, pues el almidón que se deposita
durante el día es más denso y refringente que el que se deposita durante la
noche. El aumento promedio en diámetro es de 1 micrómetro por día. La variación
en tamaño y forma del amiloplasto es característica de la especie. Por ejemplo,
en los porotos (Phaseolus spp.) los gránulos son simples y elípticos, en
las cebadas (Hordeum spp.) son esféricos de tamaño variable, en el maíz (Zea
mays) son angulosos mientras que en la avena (Avena sativa) y
el arroz (Oryza sativa) se presentan granos de almidón compuestos.
El almidón forma parte del 65-70% del peso seco de los cariópsides maduros.
Durante la germinación los granos de almidón son degradados a glucosa por
enzimas que se encuentran en la capa más externa del endosperma de las gramíneas
(capa de aleurona) y así son movilizados para su utilización durante la
germinación. Sin embargo, el almidón no es el único polisacárido de reserva.
Otros hidratos de carbono que se pueden encontrar en las semillas son las
hemicelulosas, y en en menor proporción (1-3%) se encuentran azúcares libres
como glucosa o sacarosa. La pared celular, especialmente la de las células del
pericarpo y del embrión pueden considerarse como un compartimento extra de
almacenaje de carbohidratos que posteriormente son utilizados durante la
germinación.
PROTEÍNAS
Las proteínas representan alrededor del 8-15% del peso seco de los granos de
los cereales y el 40% en las semillas de las leguminosas. Las proteínas se
almacenan como cuerpos proteicos sólidos de 0,1 a 0,25 micrómetros de diámetro,
rodeados por una membrana simple, pudiedo ser proteínas enzimáticas
(metabólicamente activas) o proteínas de reserva. Se encuentran en los
cotiledones de las leguminosas (fabáceas) y en la capa más externa del
endosperma (capa de aleurona) y en todo el endosperma de las gramíneas (poáceas)
rodeando a los gránulos de almidón y entre ellos. Los cuerpos proteicos varían
en su composición y pueden presentar inclusiones cristaloides o globoides y
cristales de oxalato de calcio como drusas. Durante la germinación las proteínas
de reserva son degradadas a aminoácidos.
Aproximadamente el 85% de las proteínas presentes en los cereales al
combinarse con el agua forman el "gluten". La propiedad visco-elástica del
gluten determina la calidad de la harina de trigo para la industria panadera,
porque otorga a la masa capacidad para retener los productos de la fermentación
de las levaduras, manteniéndose esponjosa. Por lo tanto, el parámetro más usado
por los países productores y exportadores de trigo para premiar su calidad es la
proteína. A partir de noviembre de 1995, en nuestro país se implementa un
programa de bonificaciones que beneficia a los trigos que superen el 11% de
contenido proteico. De este modo, se busca incentivar a los productores para
aumentar los niveles de calidad en la producción triguera. Por otro lado, en la
actualidad se ha comprobado que debido a cambios en los hábitos alimenticios, se
ha incrementado la demanda de bizcochos y galletas, en cuya producción
intervienen harinas que no necesitan de las propiedades panaderas. Dichas
harinas provienen de los llamados trigos blandos (de bajo contenido de
proteínas). Los principales países productores de trigo como EEUU, Canadá y
Australia producen y exportan dichos granos. Si bien los estudios de mercado
estarían indicando la conveniencia en la incorporación de trigos blandos en la
producción y exportación para nuestro país, aún su producción no está siendo
impulsada.
LÍPIDOS
Las plantas almacenan lípidos en las semillas para actuar como fuente de
energía en la germinación. Los triglicéridos constituyen los principales lípidos
de reserva en las semillas de especies oleaginosas donde se encuentran en
concentraciones excepcionalmente elevadas como grasas o aceites, contenidos en
los esferosomas de 0,2 a 6 micrómetros de diámetro. Son la forma de reserva de
energía más eficiente ya que los carbonos en los ácidos grasos (A.G.) están más
reducidos que en los hidratos de carbono; por lo tanto la oxidación de los
lípidos libera el doble de energía. El ácido graso saturado más abundante es el
palmítico. Sin embargo, los ácidos grasos insaturados oleico y linoleico (con
dobles ligaduras) se encuentran en mayor proporción y representan alrededor de
un 60% del total del aceite presente en las semillas oleaginosas. Como se
aprecia en la tabla 1, los lípidos se acumulan generalmente en los embriones,
principalmente en los cotiledones como en el caso de maní (Arachis hypogaea),
soja (Glycine max), girasol (Helianthus annuus), jojoba (Simmondsia
chinensis). Los lípidos vegetales son importantes componentes de la dieta de
humanos y animales. Además, son de particular interés para la industria de
pinturas, lubricantes y cosméticos, que constantemente demandan aceites con una
composición específica de ácidos grasos. Por esta razón se ha desarrollado un
interés en la modificación de la composición de aceites mediante aportes del
mejoramiento vegetal y la ingeniería genética, como por ejemplo el girasol alto
oleico. Recientemente se ha incorporado una nueva alternativa en el uso
industrial de los aceites vegetales, que es la producción de combustibles
ecológicos. Así nació el biodiesel que consiste en una mezcla compuesta por un
20% de aceite vegetal (de girasol, soja, colza, etc.) quimicamente tratado y un
80% de gasoil.
Un aspecto a tener en cuenta es que un inadecuado almacenamiento de la
semilla o un secado artificial incorrecto, puede desnaturalizar las proteínas
así como también perjudicar la calidad del contenido lipídico de la semilla
comprometiendo seriamente su calidad.
EMBRIÓN
El embrión se origina a partir del huevo o cigoto, y durante los estados
tempranos de su desarrollo se producen sucesivas divisiones mitóticas y
diferenciación celular. Luego de su formación, el cigoto suele entrar en un
período de reposo. La primera división del cigoto es transversal y generalmente
ocurre luego de iniciada las divisiones del núcleo endospérmico. Por lo tanto,
en el polo micropilar se diferencia una célula basal alargada, y delimitando el
polo calazal, una célula apical redondeada. La célula basal por sucesivas
divisiones forma el suspensor, que actuará en la absorción y transporte de
nutrientes hacia el embrión, el que se originará de las divisiones en diferentes
planos de la célula apical. En esta etapa, denominada histodiferenciación,
queda determinada la estructura básica de la planta.
Durante este período inicial del proceso de embriogénesis, los
nutrientes son aportados por el líquido que rodea al embrión en el saco
embrionario (nucela y endosperma). El desarrollo de los embriones de los
diversos grupos de plantas de las angiospermas tienen características comunes.
Por ejemplo, para las dicotiledóneas el embrión pasa por los estados de:
proembrión
linear, globular, embrión trapezoidal, cordiforme y torpedo. A
partir del estado trapezoidal, comienza la polarización, diferenciándose hacia
el micrópilo la radícula. Luego, en el estado cordiforme se diferencian los
cotiledones, y entre ellos el ápice caulinar. Finalmente, en el estado de
torpedo se reconocen los cotiledones, hipocótilo y radícula, figura 8.
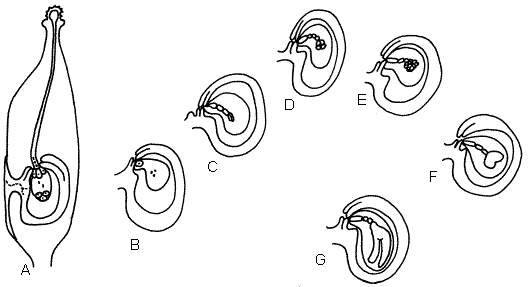 |
| Figura 8: Fecundación y
embriogénesis en una dicotiledónea. |
| A, (doble fecundación) el tubo
polínico descarga los gametos masculinos en una sinérgida: un gameto se
une con la oósfera y forma el huevo o cigoto, el otro gameto se une a los
núcleos polares y forma el núcleo endospérmico; B, huevo o cigoto (2n) y
núcleo endospérmico (3n); C, proembrión lineal; D, proembrión globular; E,
embrión trapezoidal; F, embrión cordiforme; G, embrión torpedo. |
Para el caso de las monocotiledóneas gramíneas, el desarrollo del embrión es
muy similar hasta el estado globular, a partir del cuál comienza a diferenciarse
el único cotiledón y el ápice caulinar que se presenta como una escotadura en la
base del cotiledón.
¿Cómo se produce el llenado de la semilla?
Una vez diferenciada la estructura básica, comienza la etapa de maduración
caracterizada por la expansión celular y el predominio de procesos anabólicos
que conducen a la deposición de reservas, sin aumento en el número de células.
Los asimilados requeridos en esta etapa provienen de la planta madre. Para el
caso de las leguminosas como por ejemplo la arveja (Pisum sativum),
se sabe que los fotoasimilados necesarios son aportados principalmente por los
folíolos y estípulas de la hoja axilante que lleva la rama floral y las paredes
del mismo fruto. En las gramíneas como el trigo, la fotosíntesis de la hoja
bandera
y la espiga son las que en mayor porcentaje determinan el llenado del grano. Los
nutrientes son transportados a través del haz vascular (vía simplástica) que
inerva al fruto y al óvulo a través del funículo. Sin embargo, el haz vascular
llega hasta la cálaza o a las terminaciones de sus ramificaciones, relativamente
lejos del embrión. Entonces, ¿cómo pasan los nutrientes de la generación materna
(tegumentos seminales) a la nueva generación (embrión o endosperma) si ya no hay
una conexión vascular?
Los fotoasimilados al llegar a la cálaza son descargados y bañan las paredes
celulares estableciendo una alta concentración de solutos osmóticamente activos
que, al no encontrar conexiones por la vía simplástica, pasan hacia la nueva
generación por vía apoplástica, es decir entre las paredes y los espacios
intercelulares. Además, se encuentran presentes células de transferencia con
proyecciones en sus paredes celulares, como las descriptas en el embrión de soja
(Glycine max), entre la cara abaxial de los cotiledones (en la zona de la
depresión central, concava) y la correspondiente parte interna de la cubierta
seminal con restos de endosperma (convexa), denominadas "pit y anti pit"
respectivamente. De esta forma, al aumentar la superficie de contacto con la
membrana plasmática se facilita el pasaje de nutrientes y se favorece el flujo
de solutos a través de la misma.
El éxito en el llenado del grano depende de las condiciones ambientales en el
momento en que éste comienza y durante todo el período de llenado, e involucran
complejas relaciones competitivas fuente-destino entre los órganos en el canopeo.
Por lo tanto, un déficit hídrico en éste punto es crítico y determinante del
rendimiento ya que afecta la acumulación de materia seca. La maduración se
completa cuando la semilla alcanza el máximo peso seco (madurez
fisiológica). Finalmente, en la etapa de deshidratación, se interrumpe la
conexión vascular con la planta madre, y la semilla pierde paulatinamente más
del 90% de su contenido de agua, lo que resulta en una gradual reducción del
metabolismo. A medida que el agua es eliminada de los tejidos de la semilla, el
embrión pasa a un estado metabólicamente inactivo y la semilla se convierte en
una estructura autónoma que puede permanecer en un período de latencia hasta
reiniciar la germinación.
¿Qué posición puede tener el embrión en la semilla?
El tamaño relativo del embrión es muy variable en las distintas especies. El
embrión puede ocupar todo el volumen de la semilla como en la mayoría de las
leguminosas (soja, Glycine max; garbanzo, Cicer arietinum) o ser
relativamente pequeño como en las gramíneas y ocupar la parte basal y lateral
(maíz, Zea mays; trigo (Triticum sp.).
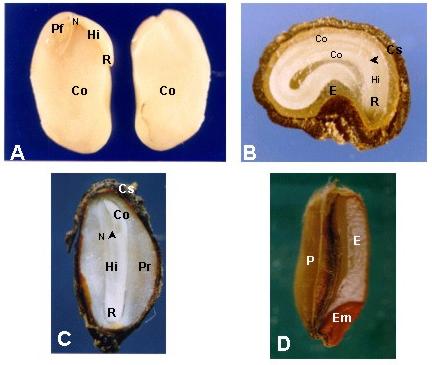 |
| Figura 9: Posición del embrión en la
semilla. |
A, embrión encorvado de soja (Glycine
max (L.) Merrill);
B, embrión curvo de chamico (Datura Ferox L.);
C, embrión axial de Pinus sp. Cs, cubierta seminal;
D, embrión lateral (Triticum sp.). Em, embrión; P,
pericarpo
Pf, primordios foliares; flecha, ápice caulinar; Co, cotiledón;
Hi, hipocótilo; N, nudo cotiledonar; E, endosperma; Pr, prótalo;
R, radícula |
Por otro lado, puede ser recto y espatulado como en el zapallo (Cucurbita
maxima) y girasol (Helianthus annuus), o curvo como en el tomate (Lycopersicon
esculentum) y pimiento (Capsicum annuum), figura 9. En la
cuscuta (Cuscuta spp.), se encuentra arrollado y en el algodón (Gossypium
hirsutum), plegado.
En los embriones curvos toma importancia la posición relativa de la radícula
y los cotiledones. De este modo, cuando la radícula se ubica frente a los bordes
de los cotiledones se denomina al embrión acumbente, si la misma se apoya
lateralmente sobre uno de los cotiledones se dice que es incumbente y
cuando además los cotiledones se pliegan longitudinalmente sobre la radícula se
denomina conduplicado, figura 10. Los embriones curvos tienen una ventaja
adaptativa ya que éstos pueden llegar a duplicar el largo de la semilla.
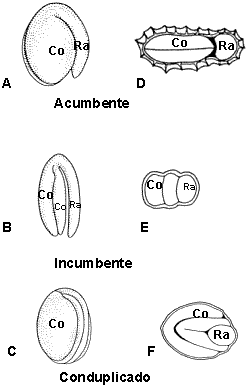 |
Figura 10: Tipos de
embriones según la ubicación de la radícula y los cotiledones.
A-C, embrión completo; D-F, corte transversal de la semilla.
A, Lepidium bonariense L. "mastuerzo loco";
B, Capsella
bursa-pastoris L. (Medikus) "bolsita del pastor";
C, Raphanus sativus
L."rabanito". |
El embrión en las gramíneas se presenta con una ubicación basal lateral
rodeado por el endosperma y la cubierta seminal o episperma soldada al
pericarpio (paredes del fruto). Según una última interpretación del embrión de
las gramíneas, la lámina del único cotiledón se encuentra dilatada a modo de
escudo formando el escutelo, la lígula de la hoja cotiledonar adopta una
configuración tubular formando el coleóptilo, que protege al ápice
caulinar y los primordios foliares. El epiblasto, presente en algunas
especies como en los trigos (Triticum spp.), representa a la vaina de la
hoja cotiledonar. Una estructura intermedia resultado de la unión entre parte de
la hoja cotiledonar e hipocótilo forma el mesocótilo. La verdadera raíz
embrional es la coleorriza que durante la germinación aborta y es
atravesada por la primera raíz adventicia, figura 11.
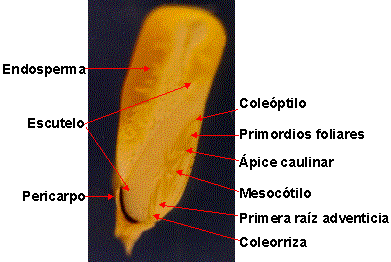 |
| Figura 11: Morfología
del cariópsis de maíz (Zea mays L.) Semilla endospermada, ubicación
del embrión basal lateral. |
En síntesis, el embrión es una planta en miniatura en estado de vida latente
o letargo con los tres órganos fundamentales de una planta adulta. La raíz está
representada por la radícula en cuyo extremo se encuentra el ápice radical, el
tallo, por el hipocótilo que termina en la gémula o plúmula que contiene el
ápice caulinar, y las hojas representadas por el o los cotiledones, según sea
una monocotiledónea, dicotiledónea o como en las gimnospermas donde los
cotiledones pueden ser más de 10.
El conocimiento de la morfología del embrión permite evaluar correctamente
las posibilidades de supervivencia de la semilla, debido a que la integridad de
cada una de sus partes es esencial ya que afectan de distinta forma la
viabilidad de la semilla. Por esta razón, es tan importante el correcto ajuste
de la maquinaria que interviene en cada una de las etapas de la producción y que
involucran la manipulación de semillas (siembra, cosecha, tratamientos post
cosecha) de modo tal que no produzcan daños mecánicos a la misma.
 Atrás
Atrás